
The Omentum in Obesity-Associated Cancer: A Hindrance to Effective Natural Killer Cell Migration towards Tumour Which Can Be Overcome by CX3CR1 Antagonism

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Demographics
2.2. Sample Preparation
2.3. Patient Sample Phenotyping
2.4. CX3CR1 Endocytosis following Treatment with Recombinant Fractalkine
2.5. CX3CR1 Recycling following Treatment with Recombinant Fractalkine
2.6. Phenotyping NK Cells following Fractalkine and IL-15 Treatment
2.7. OAC Patient-Derived Adipose Conditioned Media Treatment
2.8. NK Cell Adhesion Assay
2.9. NK Cell Chemotaxis Assay
2.10. Statistical Analysis
3. Results
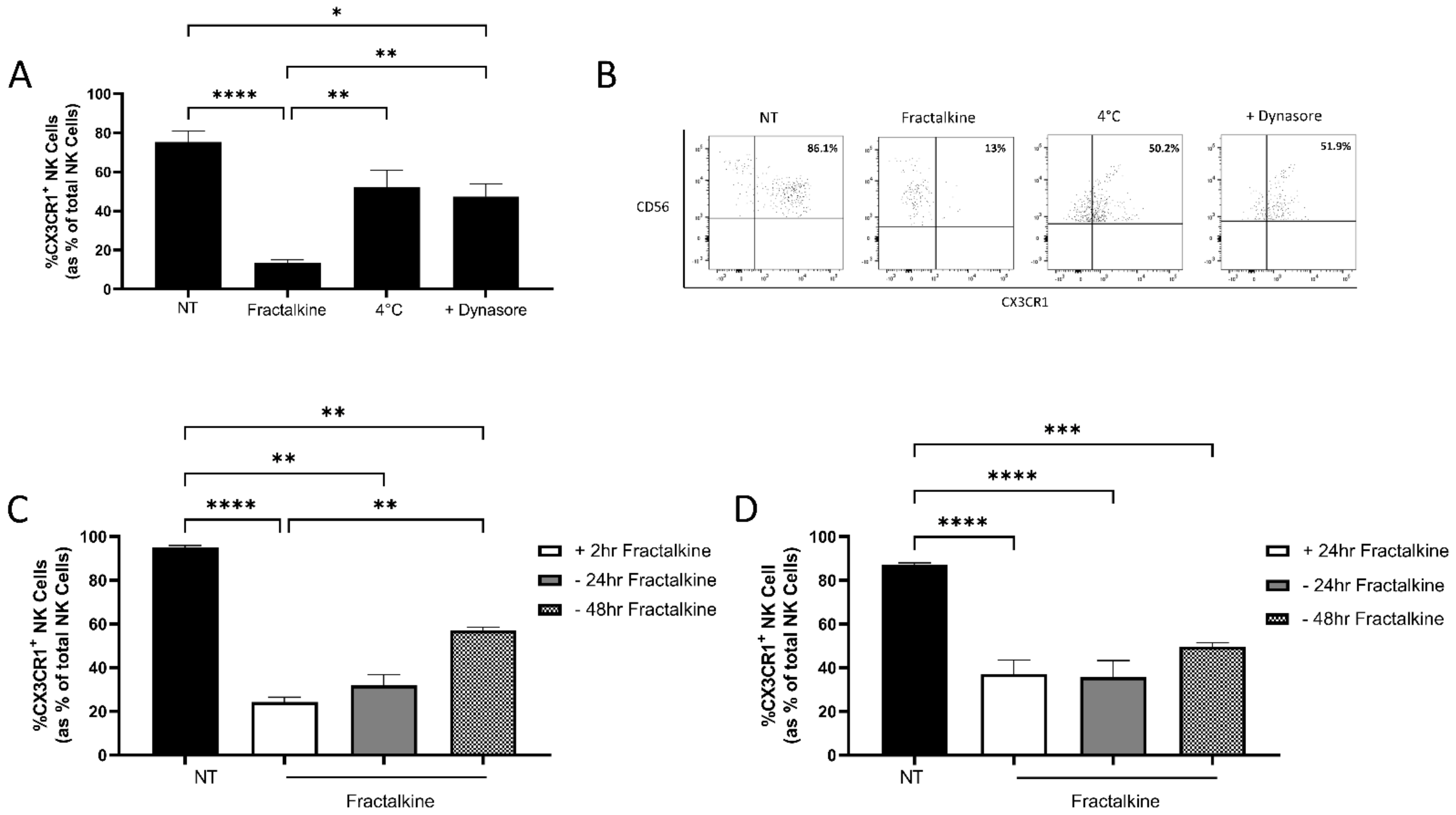
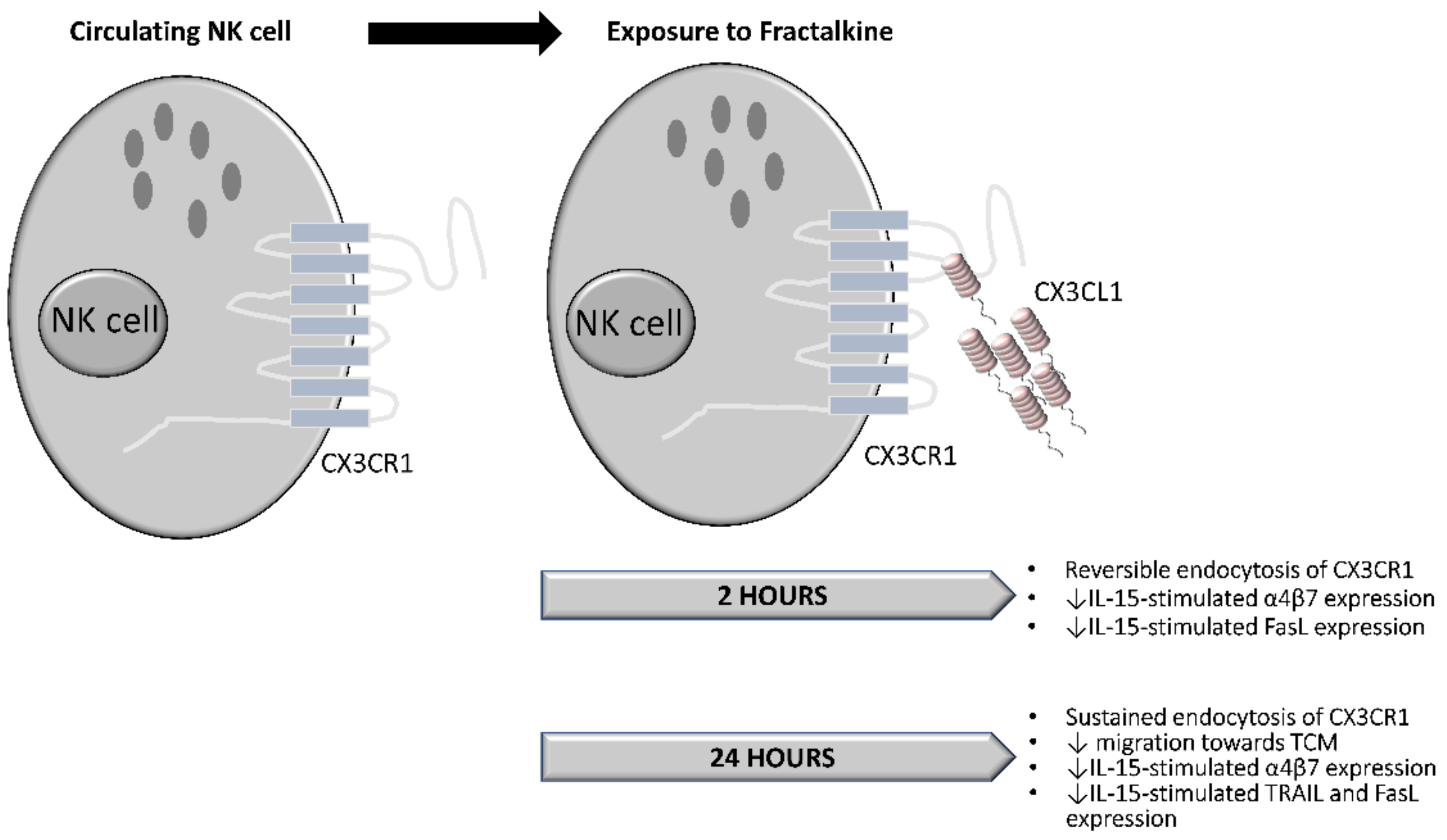
3.1. Fractalkine Mediates the Endocytosis of CX3CR1
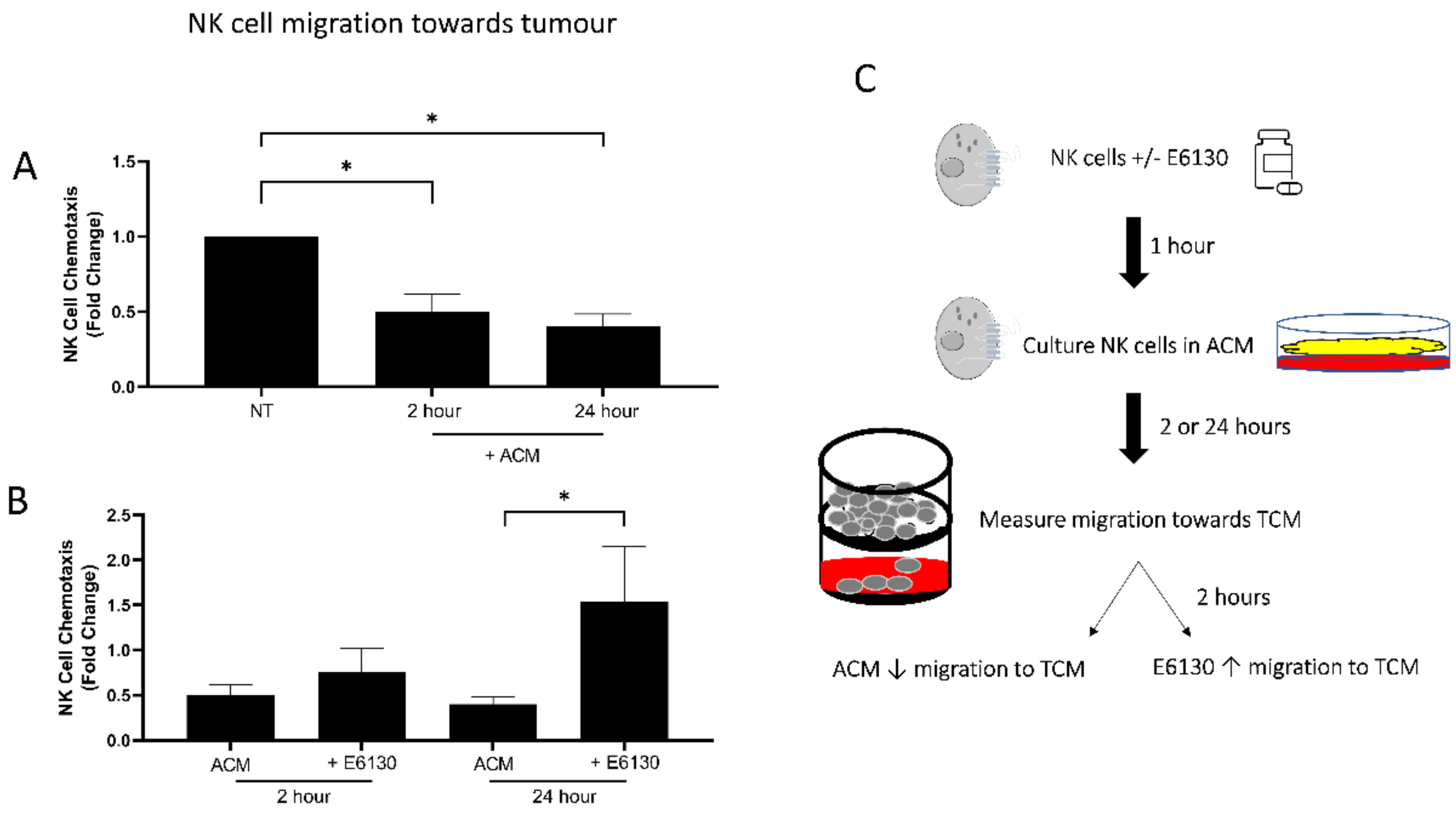
3.2. Exposure to ACM Significantly Reduces NK Cell Migration towards OAC Patient-Derived TCM
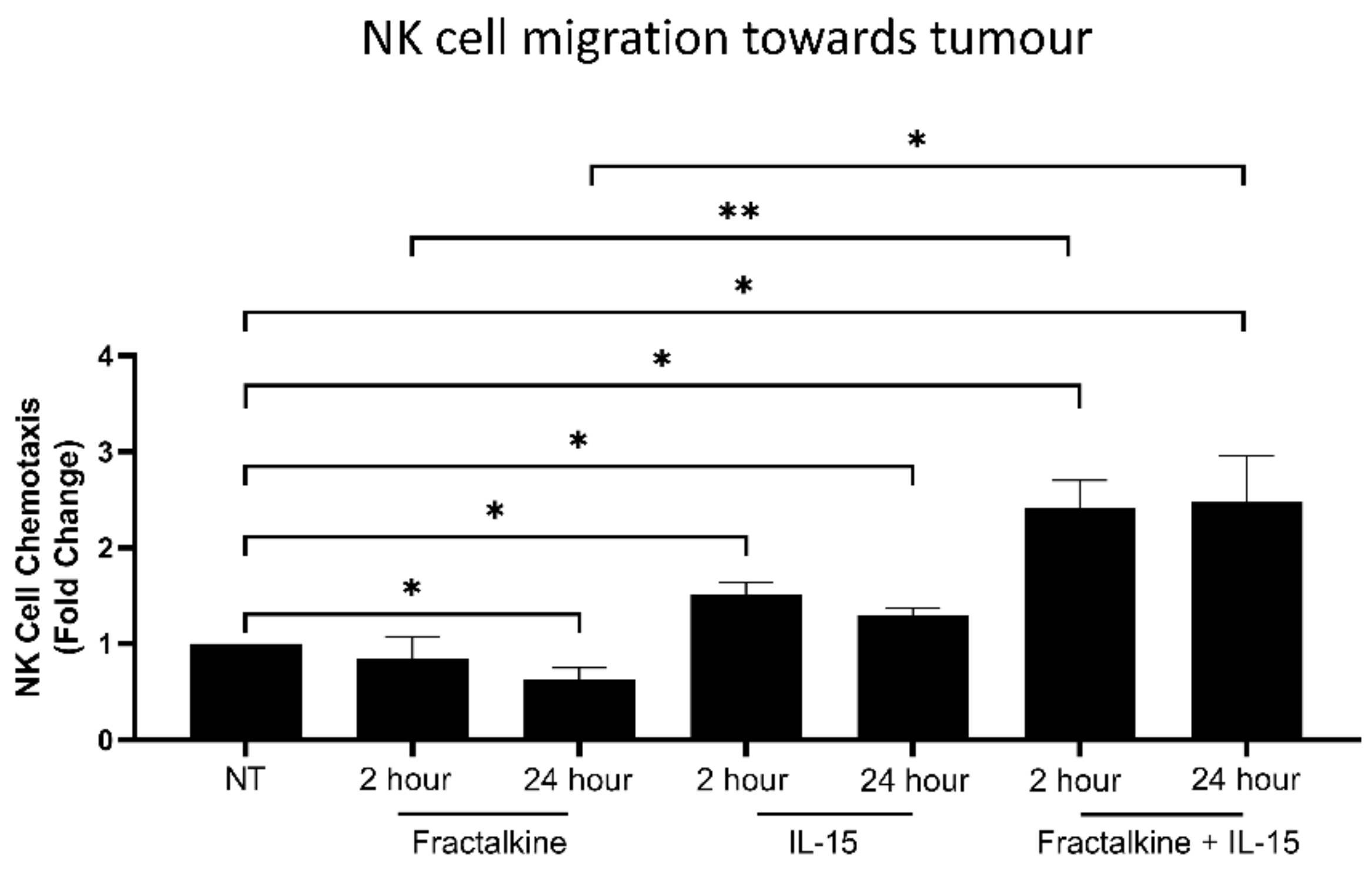
3.3. IL-15 Antagonises Fractalkine-Mediated Decreases in NK Cell Migration towards the Chemotactic Signals of OAC Tumour
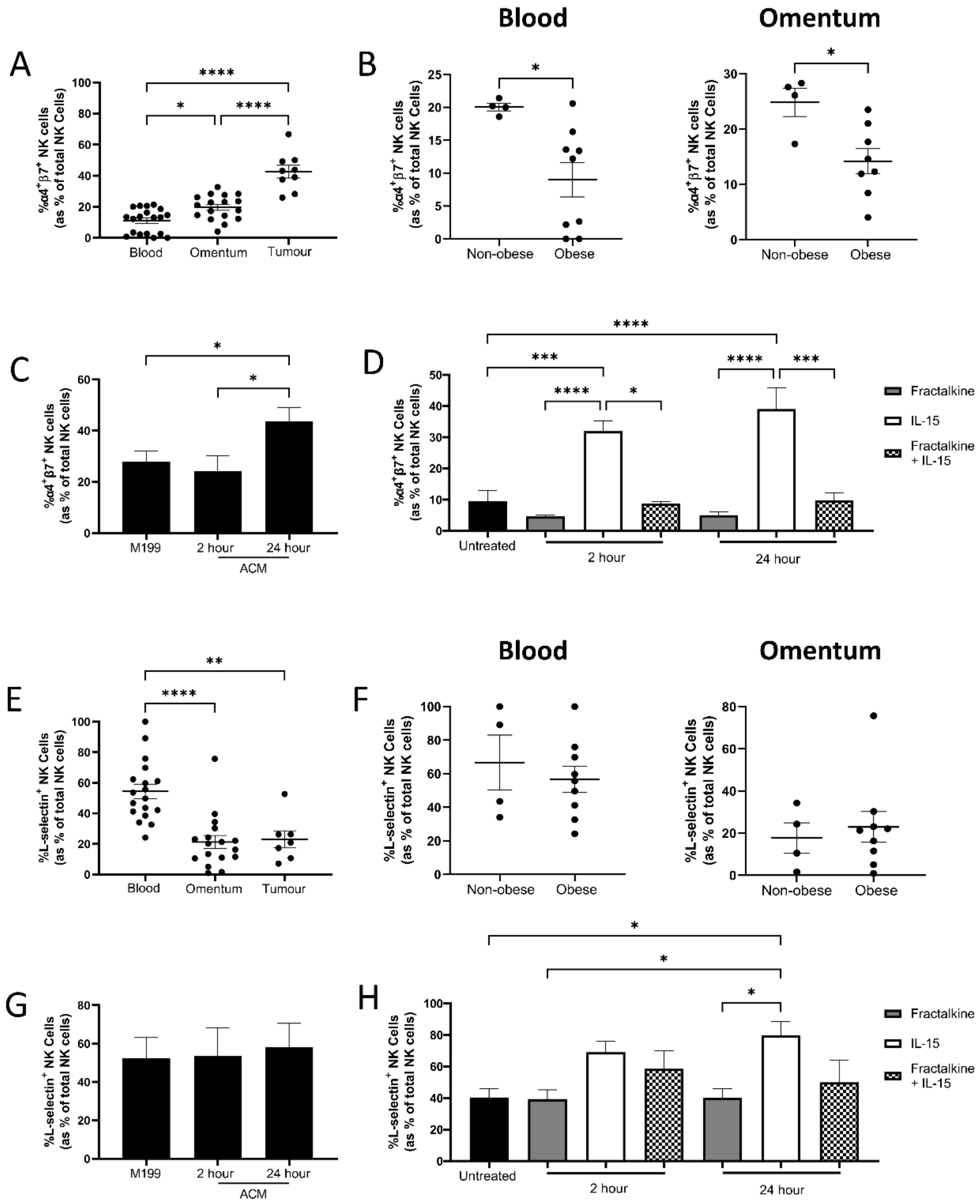
3.4. Fractalkine Antagonises IL-15-Mediated Increases in Adhesion Molecule Expression
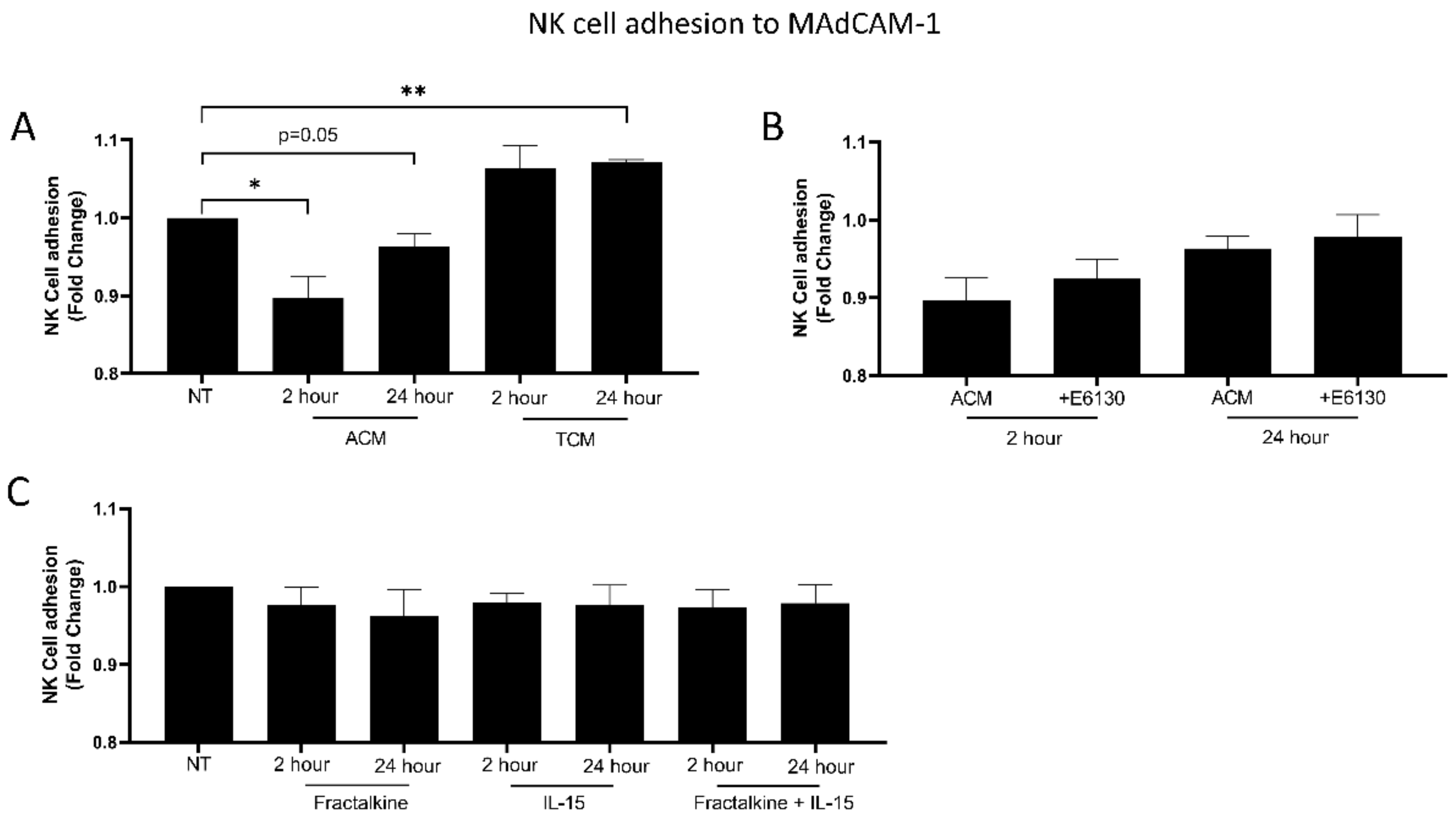
3.5. The OAC Omental Microenvironment Significantly Decreases NK Cell Adhesion to MAdCAM-1
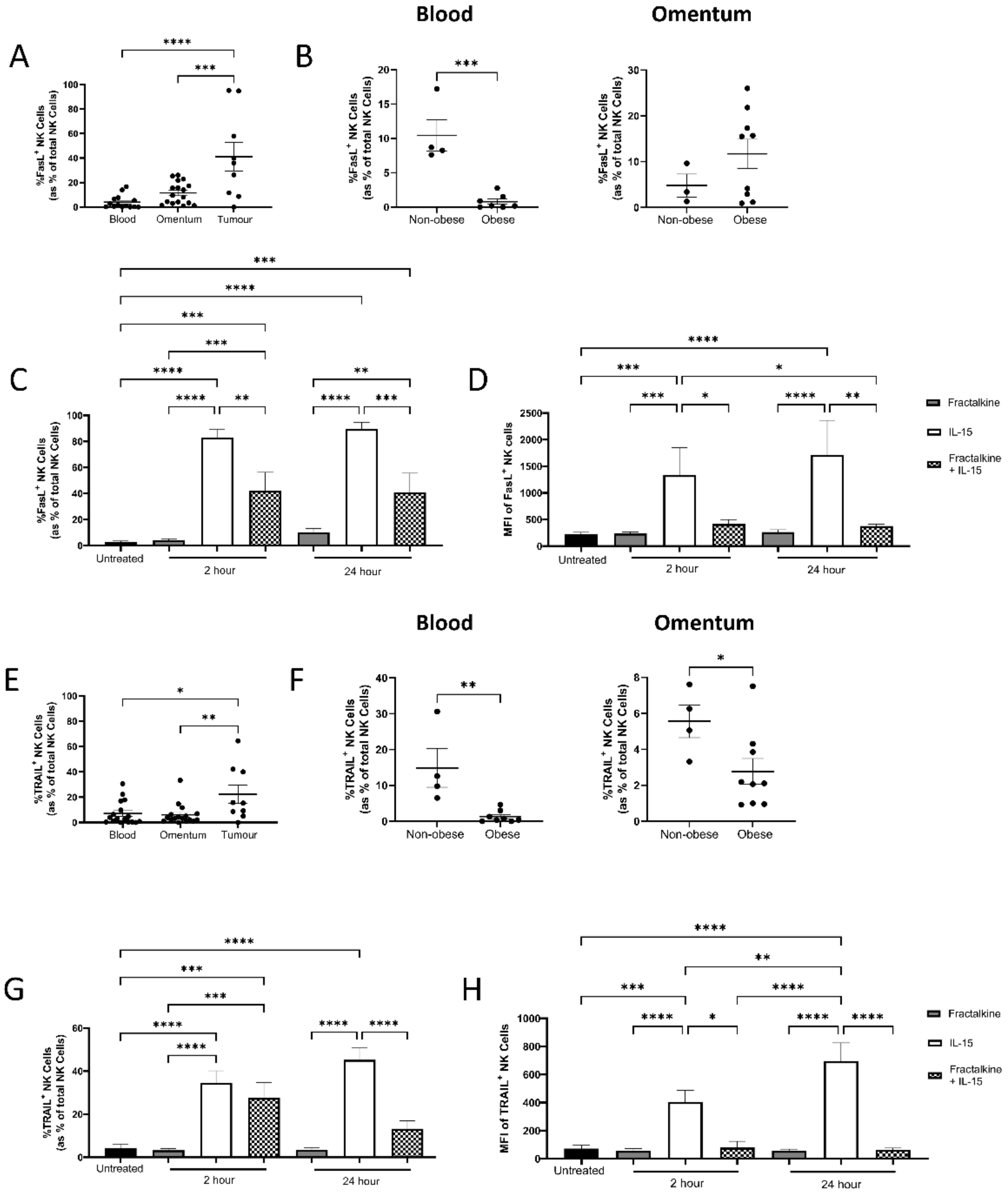
3.6. Fractalkine Significantly Reduces IL-15-Mediated Stimulation of Death Receptor Ligand Expression by NK Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef] [Green Version]
- Hoyo, C.; Cook, M.B.; Kamangar, F.; Freedman, N.D.; Whiteman, D.C.; Bernstein, L.; Brown, L.M.; Risch, H.A.; Ye, W.; Sharp, L.; et al. Body mass index in relation to oesophageal and oesophagogastric junction adenocarcinomas: A pooled analysis from the international BEACON consortium. Int. J. Epidemiol. 2012, 41, 1706–1718. [Google Scholar] [CrossRef]
- O’Sullivan, K.E.; Phelan, J.J.; O’Hanlon, C.; Lysaght, J.; O’Sullivan, J.N.; Reynolds, J.V. The role of inflammation in cancer of the esophagus. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 749–760. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA. Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Van Hagen, P.; Hulshof, M.C.C.M.; Van Lanschot, J.J.B.; Steyerberg, E.W.; Henegouwen, M.V.B.; Wijnhoven, B.P.L.; Richel, D.J.; Nieuwenhuijzen, G.A.P.; Hospers, G.A.P.; Bonenkamp, J.J.; et al. Preoperative Chemoradiotherapy for Esophageal or Junctional Cancer. N. Engl. J. Med. 2012, 366, 2074–2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ASCO, Stomach Cancer: Statistics, Cancer.Net. 2021. Available online: www.cancer.net/cancer-types/stomach-cancer/statistics (accessed on 14 October 2021).
- Ajani, J.A.; Mansfield, P.F.; Janjan, N.; Morris, J.; Pisters, P.W.; Lynch, P.M.; Feig, B.; Myerson, R.; Nivers, R.; Cohen, D.S.; et al. Multi-institutional trial of preoperative chemoradiotherapy in patients with potentially resectable gastric carcinoma. J. Clin. Oncol. 2004, 22, 2774–2780. [Google Scholar] [CrossRef] [PubMed]
- Ajani, J.A.; Mansfield, P.F.; Crane, C.H.; Wu, T.T.; Lunagomez, S.; Lynch, P.M.; Janjan, N.; Feig, B.; Faust, J.; Yao, J.C.; et al. Paclitaxel-Based Chemoradiotherapy in Localized Gastric Carcinoma: Degree of Pathologic Response and Not Clinical Parameters Dictated Patient Outcome. J. Clin. Oncol. 2005, 23, 1237–1244. [Google Scholar] [CrossRef] [PubMed]
- Vivier, E.; Tomasello, E.; Baratin, M.; Walzer, T.; Ugolini, S. Functions of natural killer cells. Nat. Immunol. 2008, 9, 503–510. [Google Scholar] [CrossRef]
- Waldhauer, I.; Steinle, A. NK cells and cancer immunosurveillance. Oncogene 2008, 27, 5932–5943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conroy, M.J.; Fitzgerald, V.; Doyle, S.L.; Channon, S.; Useckaite, Z.; Gilmartin, N.; O’Farrelly, C.; Ravi, N.; Reynolds, J.V.; Lysaght, J. The microenvironment of visceral adipose tissue and liver alter natural killer cell viability and function. J. Leukoc. Biol. 2016, 100, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Mylod, E.; Melo, A.M.; Donlon, N.E.; Davern, M.; Bhardwaj, A.; Reynolds, J.V.; Lysaght, J.; Conroy, M.J. Fractalkine Elicits Chemotactic, Phenotypic, and Functional Effects on CX3CR1 + CD27 − NK Cells in Obesity-Associated Cancer. J. Immunol. 2021, 207, 1200–1210. [Google Scholar] [CrossRef] [PubMed]
- Lin, E.W.; Karakasheva, T.A.; Hicks, P.D.; Bass, A.J.; Rustgi, A.K. The tumor microenvironment in esophageal cancer. Oncogene 2016, 35, 5337–5349. [Google Scholar] [CrossRef]
- Muro, K.; Chung, H.C.; Shankaran, V.; Geva, R.; Catenacci, D.; Gupta, S.; Eder, J.P.; Golan, T.; Le, D.T.; Burtness, B.; et al. Pembrolizumab for patients with PD-L1-positive advanced gastric cancer (KEYNOTE-012): A multicentre, open-label, phase 1b trial. Lancet Oncol. 2016, 17, 717–726. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. Mechanisms of disease: The many roles of chemokines and chemokine receptors in inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, C.; Davern, M.; Donlon, N.E.; Lysaght, J.; Conroy, M.J. Chemokine-targeted therapies: An opportunity to remodel immune profiles in gastro-oesophageal tumours. Cancer Lett. 2021, 521, 224–236. [Google Scholar] [CrossRef]
- Conroy, M.J.; Maher, S.G.; Melo, A.M.; Doyle, S.L.; Foley, E.; Reynolds, J.V.; Long, A.; Lysaght, J. Identifying a novel role for fractalkine (CX3CL1) in memory CD8+ T cell accumulation in the omentum of obesity-associated cancer patients. Front. Immunol. 2018, 9, 1867. [Google Scholar] [CrossRef] [Green Version]
- Polyák, A.; Ferenczi, S.; Dénes, A.; Winkler, Z.; Kriszt, R.; Pintér-Kübler, B.; Kovács, K.J. The fractalkine/Cx3CR1 system is implicated in the development of metabolic visceral adipose tissue inflammation in obesity. Brain Behav Immun. 2014, 38, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.; Hinkle, C.C.; Ferguson, J.F.; Mehta, N.N.; Li, M.; Qu, L.; Lu, Y.; Putt, M.E.; Ahima, R.S.; Reilly, M.P. Fractalkine is a novel human adipochemokine associated with type 2 diabetes. Diabetes 2011, 60, 1512–1518. [Google Scholar] [CrossRef] [Green Version]
- D’Haese, J.G.; Demir, I.E.; Friess, H.; Ceyhan, G.O. Fractalkine/CX3CR1: Why a single chemokine-receptor duo bears a major and unique therapeutic potential. Expert Opin. Ther. Targets 2010, 14, 207–219. [Google Scholar] [CrossRef]
- Conroy, M.J.; Lysaght, J. CX3CL1 Signaling in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1231, 1–12. [Google Scholar] [CrossRef]
- Waldmann, T.A. Interleukin-15 in the treatment of cancer. Expert Rev. Clin. Immunol. 2014, 10, 1689–1701. [Google Scholar] [CrossRef]
- Waldmann, T.A. The biology of interleukin-2 and interleukin-15: Implications for cancer therapy and vaccine design. Nat. Rev. Immunol. 2006, 6, 595–601. [Google Scholar] [CrossRef]
- Yang, Y.; Lundqvist, A. Immunomodulatory Effects of IL-2 and IL-15; Implications for Cancer Immunotherapy. Cancers 2020, 12, 3586. [Google Scholar] [CrossRef]
- Lysaght, J.; Allott, E.H.; Donohoe, C.L.; Howard, J.M.; Pidgeon, G.P.; Reynolds, J.V. T lymphocyte activation in visceral adipose tissue of patients with oesophageal adenocarcinoma. Br. J. Surg. 2011, 98, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.M.; O’Brien, A.M.; Phelan, J.J.; Kennedy, S.A.; Wood, N.A.W.; Veerapen, N.; Besra, G.S.; Clarke, N.E.; Foley, E.K.; Ravi, A.; et al. Mucosal-Associated Invariant T Cells Display Diminished Effector Capacity in Oesophageal Adenocarcinoma. Front. Immunol. 2019, 10, 1580. [Google Scholar] [CrossRef] [PubMed]
- Lysaght, J.; van der Stok, E.P.; Allott, E.H.; Casey, R.; Donohoe, C.L.; Howard, J.M.; McGarrigle, S.A.; Ravi, N.; Reynolds, J.V.; Pidgeon, G.P. Pro-inflammatory and tumour proliferative properties of excess visceral adipose tissue. Cancer Lett. 2011, 312, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Strazza, M.; Azoulay-Alfaguter, I.; Pedoeem, A.; Mor, A. Static Adhesion Assay for the Study of Integrin Activation in T Lymphocytes. J. Vis. Exp. 2014, 88, 51646. [Google Scholar] [CrossRef] [Green Version]
- Wagner, N.; Löhler, J.; Kunkel, E.J.; Ley, K.; Leung, E.; Krissansen, G.; Rajewsky, K.; Müller, W. Critical role for beta7 integrins in formation of the gut-associated lymphoid tissue. Nature 1996, 382, 366–370. [Google Scholar] [CrossRef]
- Briskin, M.; Winsor-Hines, D.; Shyjan, A.; Cochran, N.; Bloom, S.; Wilson, J.; McEvoy, L.M.; Butcher, E.C.; Kassam, N.; Mackay, C.R.; et al. Human mucosal addressin cell adhesion molecule-1 is preferentially expressed in intestinal tract and associated lymphoid tissue. Am. J. Pathol. 1997, 151, 97–110. [Google Scholar]
- Smyth, M.J.; Cretney, E.; Kelly, J.M.; Westwood, J.A.; Street, S.E.A.; Yagita, H.; Takeda, K.; Dommelen, S.L.H.V.; Degli-Esposti, M.A.; Hayakawa, Y. Activation of NK cell cytotoxicity. Mol. Immunol. 2005, 42, 501–510. [Google Scholar] [CrossRef]
- Ishigami, S.; Natsugoe, S.; Tokuda, K.; Nakajo, A.; Che, X.; Iwashige, H.; Aridome, K.; Hokita, S.; Aikou, T. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer 2000, 88, 577–583. [Google Scholar] [CrossRef]
- Xu, B.; Chen, L.; Li, J.; Zheng, X.; Shi, L.; Wu, C.; Jiang, J. Prognostic value of tumor infiltrating NK cells and macrophages in stage II+III esophageal cancer patients. Oncotarget 2016, 7, 74904–74916. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Berger, M.; Hackman, R.C.; Gough, M.; Elliott, C.; Jensen, M.C.; Riddell, S.R. Safety and immunologic effects of IL-15 administration in nonhuman primates. Blood 2009, 114, 2417–2426. [Google Scholar] [CrossRef] [Green Version]
- Allavena, P.; Giardina, G.; Bianchi, G.; Mantovani, A. IL-15 is chemotactic for natural killer cells and stimulates their adhesion to vascular endothelium. J. Leukoc. Biol. 1997, 61, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Nayyar, G.; Chu, Y.; Cairo, M.S. Overcoming resistance to natural killer cell based immunotherapies for solid tumors. Front. Oncol. 2019, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, J.; Niu, J.; Zhang, J.; Tian, Z. Interleukin-15 improves cytotoxicity of natural killer cells via up-regulating NKG2D and cytotoxic effector molecule expression as well as STAT1 and ERK1/2 phosphorylation. Cytokine 2008, 42, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Villar, J.J.; Zapata, J.M.; Melero, I.; Postigo, A.; Sánchez-Madrid, E.; López-Botet, M. Expression and function of alpha 4/beta 7 integrin on human natural killer cells. Immunology 1996, 89, 96–104. [Google Scholar] [CrossRef]
- Connor, E.M.; Eppihimer, M.J.; Morise, Z.; Granger, D.N.; Grisham, M.B. Expression of mucosal addressin cell adhesion molecule-1 (MAdCAM-1) in acute and chronic inflammation. J. Leukoc. Biol. 1999, 65, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Berberich, S.; Dähne, S.; Schippers, A.; Peters, T.; Müller, W.; Kremmer, E.; Förster, R.; Pabst, O. Differential molecular and anatomical basis for B cell migration into the peritoneal cavity and omental milky spots. J. Immunol. 2008, 180, 2196–2203. [Google Scholar] [CrossRef] [Green Version]
- Carlow, D.A.; Gold, M.R.; Ziltener, H.J. Lymphocytes in the peritoneum home to the omentum and are activated by resident dendritic cells. J. Immunol. 2009, 183, 1155–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meza-Perez, S.; Randall, T.D. Immunological Functions of the Omentum. Trends Immunol. 2017, 38, 526–536. [Google Scholar] [CrossRef]
- Salmi, M.; Andrew, D.P.; Butcher, E.C.; Jalkanen, S. Dual binding capacity of mucosal immunoblasts to mucosal and synovial endothelium in humans: Dissection of the molecular mechanisms. J. Exp. Med. 1995, 181, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Kavanagh, M.E.; Conroy, M.J.; Clarke, N.E.; Gilmartin, N.T.; O’Sullivan, K.E.; Feighery, R.; MacCarthy, F.; O’Toole, D.; Ravi, N.; Reynolds, J.V.; et al. Impact of the inflammatory microenvironment on T-cell phenotype in the progression from reflux oesophagitis to Barrett oesophagus and oesophageal adenocarcinoma. Cancer Lett. 2016, 370, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.-J.; Chen, J.-Y.; Kuo, M.-L.; Hsiao, H.-S.; Lee, P.-T.; Huang, J.-L. Effect of Interleukin-15 on CD11b, CD54, and CD62L Expression on Natural Killer Cell and Natural Killer T-Like Cells in Systemic Lupus Erythematosus. Mediators Inflamm. 2016, 2016, 9675861. [Google Scholar] [CrossRef]
- Mishra, H.K.; Dixon, K.J.; Pore, N.; Felices, M.; Miller, J.S.; Walcheck, B. Activation of ADAM17 by IL-15 Limits Human NK Cell Proliferation. Front. Immunol. 2021, 12, 1621. [Google Scholar] [CrossRef]
- Yamada, M.; Yanaba, K.; Hasegawa, M.; Matsushita, Y.; Horikawa, M.; Komura, K.; Matsushita, T.; Kawasuji, A.; Fujita, T.; Takehara, K.; et al. Regulation of local and metastatic host-mediated anti-tumour mechanisms by l-selectin and intercellular adhesion molecule-1. Clin. Exp. Immunol. 2006, 143, 216–227. [Google Scholar] [CrossRef]
- Berg, E.L.; McEvoy, L.M.; Berlin, C.; Bargatze, R.F.; Butcher, E.C. L-selectin-mediated lymphocyte rolling on MAdCAM-1. Nature 1993, 366, 695–698. [Google Scholar] [CrossRef]
- Sobolev, O.; Stern, P.; Lacy-Hulbert, A.; Hynes, R.O. Natural killer cells require selectins for suppression of subcutaneous tumors. Cancer Res. 2009, 69, 2531–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Kawashima, H.; Lowe, J.B.; Lanier, L.L.; Fukuda, M. Suppression of tumor formation in lymph nodes by L-selectin-mediated natural killer cell recruitment. J. Exp. Med. 2005, 202, 1679–1689. [Google Scholar] [CrossRef]
- Juelke, K.; Killig, M.; Luetke-Eversloh, M.; Parente, E.; Gruen, J.; Morandi, B.; Ferlazzo, G.; Thiel, A.; Schmitt-Knosalla, I.; Romagnani, C. CD62L expression identifies a unique subset of polyfunctional CD56 dim NK cells. Blood 2010, 116, 1299–1307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harjunpää, H.; Llort Asens, M.; Guenther, C.; Fagerholm, S.C. Cell Adhesion Molecules and Their Roles and Regulation in the Immune and Tumor Microenvironment. Front. Immunol. 2019, 10, 78. [Google Scholar] [CrossRef] [Green Version]
- Cibrián, D.; Sánchez-Madrid, F. CD69: From activation marker to metabolic gatekeeper. Eur. J. Immunol. 2017, 47, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Zamai, L.; Ahmad, M.; Bennett, I.M.; Azzoni, L.; Alnemri, E.S.; Perussia, B. Natural Killer (NK) Cell–mediated Cytotoxicity: Differential Use of TRAIL and Fas Ligand by Immature and Mature Primary Human NK Cells. J. Exp. Med. 1998, 188, 2375–2380. [Google Scholar] [CrossRef]
- Mirandola, P.; Ponti, C.; Gobbi, G.; Sponzilli, I.; Vaccarezza, M.; Cocco, L.; Zauli, G.; Secchiero, P.; Manzoli, F.A.; Vitale, M. Activated human NK and CD8+ T cells express both TNF-related apoptosis-inducing ligand (TRAIL) and TRAIL receptors but are resistant to TRAIL-mediated cytotoxicity. Blood 2004, 104, 2418–2424. [Google Scholar] [CrossRef] [PubMed]
- Peter, M.E.; Hadji, A.; Murmann, A.E.; Brockway, S.; Putzbach, W.; Pattanayak, A.; Ceppi, P. The role of CD95 and CD95 ligand in cancer. Cell Death Differ. 2015, 22, 549–559. [Google Scholar] [CrossRef] [PubMed]
- O’Shea, D.; Hogan, A.E. Dysregulation of Natural Killer Cells in Obesity. Cancers 2019, 11, 573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bähr, I.; Spielmann, J.; Quandt, D.; Kielstein, H. Obesity-Associated Alterations of Natural Killer Cells and Immunosurveillance of Cancer. Front. Immunol. 2020, 11, 245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laue, T.; Wrann, C.D.; Hoffmann-Castendiek, B.; Pietsch, D.; Hübner, L.; Kielstein, H. Altered NK cell function in obese healthy humans. BMC Obes. 2015, 2, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (years) | 64 |
| Sex ratio (M:F) | 20:8 |
| Diagnosis (no. patients) | |
| OAC | 9 |
| OGJ | 14 |
| Gastric | 5 |
| Tumour Stage a (no. patients) | |
| T0 | 3 |
| T1 | 6 |
| T2 | 7 |
| T3 | 6 |
| T4 | 5 |
| Nodal Status b (no. patients) | |
| Positive | 12 |
| Negative | 13 |
| Mean BMI (kg/m2) | 29 |
| BMI c (no. patients) | |
| Underweight (BMI < 19.9) | 0 |
| Normal Weight (BMI 20–24.9) | 4 |
| Overweight (BMI 25–29.9) | 12 |
| Obese (BMI > 30) | 11 |
| Mean VFA (cm2) d | 144 |
| Viscerally Obese by VFA e | 60% |
| Received Neoadjuvant CRT | 68% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mylod, E.; O’Connell, F.; Donlon, N.E.; Butler, C.; Reynolds, J.V.; Lysaght, J.; Conroy, M.J. The Omentum in Obesity-Associated Cancer: A Hindrance to Effective Natural Killer Cell Migration towards Tumour Which Can Be Overcome by CX3CR1 Antagonism. Cancers 2022, 14, 64. https://doi.org/10.3390/cancers14010064
Mylod E, O’Connell F, Donlon NE, Butler C, Reynolds JV, Lysaght J, Conroy MJ. The Omentum in Obesity-Associated Cancer: A Hindrance to Effective Natural Killer Cell Migration towards Tumour Which Can Be Overcome by CX3CR1 Antagonism. Cancers. 2022; 14(1):64. https://doi.org/10.3390/cancers14010064
Chicago/Turabian StyleMylod, Eimear, Fiona O’Connell, Noel E. Donlon, Christine Butler, John V. Reynolds, Joanne Lysaght, and Melissa J. Conroy. 2022. "The Omentum in Obesity-Associated Cancer: A Hindrance to Effective Natural Killer Cell Migration towards Tumour Which Can Be Overcome by CX3CR1 Antagonism" Cancers 14, no. 1: 64. https://doi.org/10.3390/cancers14010064